



Seymouriamorpha
Michel Laurin



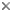
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Seymouriamorphs are a small but widespread group of early terrestrial choanates. They are known from the Lower to Upper Permian and have been found in Europe, Asia, and North America (Fig. 1). Six genera and eight species are known. Many additional species have been erected, but most of them are not valid (Klembara and Janiga, 1993).
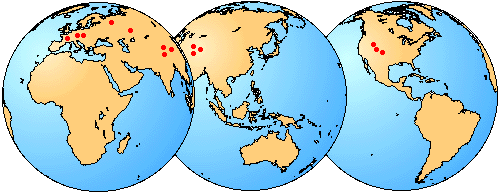
Figure 1. Geographic distribution of seymouriamorph localities. These localities are located in Eurasia and North America, but in the Permian, the continents were in different positions and Eurasia was represented by several smaller plates.
Small aquatic larvae bearing external gills are known in some seymouriamorphs (Ivakhnenko, 1981; Kuznetsov and Ivakhnenko, 1981; Klembara, 1995). The adults, when known, appear to have been terrestrial. The skull length of the known specimens ranges from about 6 mm in the smallest larvea (Ivakhnenko, 1981) to 15 cm in presumably mature individuals. The monophyly of this group has long been doubted because of size and ontogenetic differences between various taxa. Seymouria and Kotlassia are only known from relatively large and mature specimens (Fig. 2) whereas all other known seymouriamorphs are represented by larval and small to mid-sized postmetamorphic specimens and were previously called discosauriscids (this is now known to be a paraphyletic group). However, recent discoveries of fairly large specimens of Discosauriscus (Klembara and Meszáros, 1992) and Ariekanerpeton (Laurin, 1996b) and relatively small specimens of Seymouria (Berman and Martens, 1993) have bridged this gap.
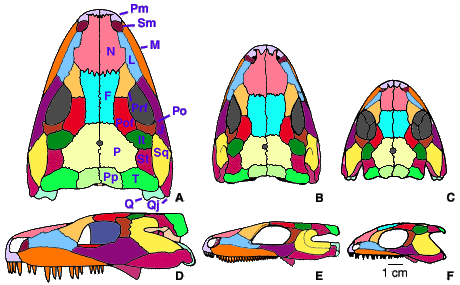
Figure 2. Seymouriamorph skulls in dorsal (A-C) and left lateral (D-F) views. Seymouria baylorensis (A, D). Seymouria sanjuanensis (B, E). Ariekanerpeton sigalovi (E, F). The specimens on which the reconstructions of S. sanjuanensis and A. sigalovi are based are probably not mature and these taxa may have reached approximately the same size as S. baylorensis. Modified from Laurin (1995).
Characteristics
The gut contents or coprolites of seymouriamorphs have only been found recently (Klembaraand Meszáros, 1992), and they suggest that seymouriamorphs resorted at least occasionally to cannibalism. Their main diet is unknown. However, their sharp, conical teeth and palatal fangs suggest that seymouriamorphs were predators.
The dermal cranial bones of adult seymouriamorphs are deeply sculptured and exhibit a hexagonal pattern of ridges also found in many temnospondyls (Fig. 3). The larvae lacked this sculpturing but they were covered by shallow pits and grooves radiating from the center of ossification of the bones. The presence of dermal sculpturing suggests that the skin was tightly attached to the skull. Seymouriamorphs retained all the dermal bones found in most other groups of terrestrial choanates, and, as in embolomeres and amniotes, the tabular contacted the parietal.

Figure 3. Dorsal view of the skull of a juvenile postmetamorphic specimen of Discosauriscus austriacus showing the dermal sculpturing that characterizes relatively mature seymouriamorphs. Courtesy of Dr. Jozef Klembara.
Seymouriamorphs had a relatively short trunk (24 to 28 presacral vertebrae) and stout limbs. Each vertebra was composed of a neural arch, a large, cylindrical, amphicoelous pleurocentrum, and a small, crescentic intercentrum. The neural arches were swollen, especially in large, mature specimens. The atlantal (first cervical) arch and pleurocentrum were paired. This is probably a primitive character because the neural arches and pleurocentra of the earliest terrestrial vertebrates were also paired (they had independent left and right arches and pleurocentra rather than a fused median arch and pleurocentrum). Hemal arches were present in the tail. There was usually only one sacral vertebra in small specimens, but large individuals occasionally had two. In this case, the posterior pair of sacral ribs was much more slender than the anterior pair, as in Seymouria baylorensis, or only one of the anterior ribs (right or left) contacted the ilium, as in Kotlassia. Ribs were present from the first cervical to the first few caudal vertebrae. The ribs of the pectoral region were expanded, presumably to provide better attachment to the muscles that supported the shoulder girdle. The exact number of caudal ribs is unknown, but as many as fifteen pairs may have been present (Ivakhnenko, 1981).
The dermal shoulder girdle included a median interclavicle and a paired clavicle and cleithra. The endochondral shoulder girdle included a scapula and a coracoid. Like many other terrestrial choanates, seymouriamorphs had five fingers and toes. The phalangeal formula was 2 3 4 4 3 or 2 3 4 5 3 in the hand (depending on the genus) and 2 3 4 5 3 in the foot.
Seymouriamorphs share a few derived characters, but an exhaustive list is difficult to establish because most taxa are incompletely known. However, all seymouriamorphs have relatively small posttemporal fenestrae (these are openings on the occiput through which blood vessels enter the skull), although this character is especially noticeable in large, mature seymouriamorphs. Their posttemporal fenestra may have grown with a negative allometry.
All seymouriamorphs appear to have an otic tube composed of the opisthotic, the prootic, and the parasphenoid.
Most, if not all, seymouriamorphs had a large, transverse dorsal flange of the pterygoid (called the lamina ascendens by Bystrow, 1944) that extended to the skull roof.
All seymouriamorphs had a slender stapes that may have been involved in hearing. This character appeared convergently in temnospondyls and in several groups of tetrapods.
Discussion of Phylogenetic Relationships
Seymouriamorphs have long played a prominent role in scenarios and theories on the origin of amniotes. They were originally believed to be closely related to amniotes or even to represent the most archaic amniotes (Broili, 1904, White, 1939). The discovery of larval seymouriamorphs with external gills contradicts this hypothesis. However, many scientists still consider them close relatives of amniotes (Gauthier et al., 1988). Despite their long association with the origin of amniotes, seymouriamorphs have long been considered amphibians simplybecause they are not amniotes, and all anamniotic terrestrial vertebrates were considered amphibians before classifications were modified to include only monophyletic groups (Carroll, 1988). However, seymouriamorphs are not closely related to lissamphibians and are not amphibians in the modern sense of this word.
Some recent studies suggesting that seymouriamorphs are not as closely related to amniotes as previously thought also assert that lepospondyls were more closely related to amniotes than to seymouriamorphs (Carroll, 1995). Others (Laurin and Reisz, 1997) argued that seymouriamorphs were not more closely related to amniotes than to amphibians. If the latter view is correct, seymouriamorphs are not tetrapods because Tetrapoda is defined as the crown-group of terrestrial choanates (Gauthier et al., 1989).
Few studies have dealt with the phylogeny within Seymouriamorpha. However, recent work has suggested that Discosauriscus, Ariekanerpeton, and Seymouria are more closely related to each other than to Kotlassia and Utegenia (Laurin, in press). The affinities of Urumqia are difficult to evaluate because this seymouriamorph is poorly known (Zhang et al., 1984), but the presence of rhomboidal ventral bony scales (gastralia) suggests that it is not closely related to Discosauriscus, Ariekanerpeton, and Seymouria.
Sensory Organs
The dermal cranial bones of larval and perhaps some juvenile postmetamorphic specimens of seymouriamorphs bear grooves for the lateral-line organ. This organ is composed of ciliated cells that can detect motion in the water. Only aquatic vertebrates have a lateral-line. The lateral-line organ is found in all fishes, most permanently aquatic amphibians, and many aquatic larvae of terrestrial amphibians. Therefore, juvenile seymouriamorphs were certainly aquatic. However, the absence of grooves in large postmetamorphic specimens of seymouriamorphs suggests the lateral-line may have disappeared during or after the metamorphosis.
Enigmatic structures consisting of foramina located in deep depressions were recently discovered on some cranial bones of larval specimens of Discosauriscus (Klembara, 1994). These structures have been interpreted as foraminate pits, structures that were previously known in osteolepiforms, but in no terrestrial vertebrate. Comparisons with urodeles suggest that the foraminate pits housed ampullary electroreceptive organs. Similar electroreceptive organs are widespread in vertebrates and are found, among other groups, in sharks, teleosts, and lungfishes. However, in all extant taxa, the ampullary organ is located in soft tissues and leaves no trace on the skeleton. Therefore, it is difficult to prove that the foraminate pits of Discosauriscus housed ampullary organs.
The slender stapes of seymouriamorphs and the presence of a large otic notch in the temporal area suggest that seymouriamorphs had a tympanic middle ear. The otic notch probably supported a large tympanum (ear drum) and the stapes probably transmitted the sounds from the tympanum to the fenestra ovalis (the stapes is involved in sound transmission in all tetrapods that have a tympanum, although in mammals, other bones are also involved). The tympanic middle ear is an adaptation to hear high-frequency air-borne sounds (it is not effective underwater). Tetrapods lacking a tympanum, like salamanders, can detect low-frequency seismic vibrations (Duellman and Trueb, 1986), but they cannot detect high-frequency air-borne sounds (above 1000 Hz). The presence of a tympanic middle ear in seymouriamorphs and the absence of lateral-line canal grooves in large postmetamorphic specimens suggest that adult seymouriamorphs were terrestrial.
References
Berman, D. S. & T. Martens. 1993. First occurrence of Seymouria (Amphibia: Batrachosauria) in the Lower Permian Rotliegend of central Germany. Annals of Carnegie Museum 62: 63-79.
Broili, F. 1904. Stammreptilien. Anatomischer Anzeiger 25: 577-587.
Bystrow A. P. 1944. Kotlassia prima Amalitzky. Geological Society of America Bulletin 55: 379-416.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution. New York: W. H. Freeman and Company.
Carroll, R. L. 1995. Problems of the phylogenetic analysis of Paleozoic choanates. Bulletin du Muséum national d'Histoire naturelle de Paris 4ème série 17: 389-445.
Duellman, W. E. & L. Trueb. 1986. Biology of Amphibians. New York: McGraw-Hill.
Gauthier, J., A. G. Kluge, & T. Rowe. 1988. The early evolution of the Amniota. In M. J. Benton (ed.) The phylogeny and classification of the tetrapods, Volume 1: amphibians, reptiles, birds: 103-155. Oxford: Clarendon Press.
Gauthier, J., D. C. Cannatella, K. De Queiroz, A. G. Kluge, & T. Rowe. 1989. Tetrapod phylogeny. In B. Fernholm, K. Bremer, and H. Jornvall (eds.) The Hierarchy of Life: 337-353. New York: Elsevier Science Publishers B. V. (Biomedical Division).
Ivakhnenko, M. F. 1981. Discosauriscidae from the Permian of Tadzhikistan. Paleontological Journal 1981: 90-102.
Klembara, J. 1994. Electroreceptors in the Lower Permian tetrapod Discosauriscus austriacus. Palaeontology 37: 609-626.
Klembara, J. 1995. The external gills and ornamentation of skull roof bones of the Lower Permian tetrapod Discosauriscus (Kuhn 1933) with remarks to its ontogeny. Paläontologische Zeitschrift 69: 265-281.
Klembara, J. & S. Meszáros. 1992. New finds of Discosauriscus austriacus (Makowsky 1876) from the Lower Permian of Boskovice furrow (Czecho-Slovakia). Geologica Carpathica 43: 305-312.
Klembara, J. & M. Janiga. 1993. Variation in Discosauriscus austriacus (Makowsky 1876) from the Lower Permian of the Boskovice furrow (Czech Republic). Zoological Journal of the Linnean Society 108: 247-270.
Kuznetsov, V. V. & M. F. Ivakhnenko. 1981. Discosauriscids from the Upper Paleozoic in Southern Kazakhstan. Paleontological Journal 1981: 101-108.
Laurin, M. 1995. Comparative cranial anatomy of Seymouria sanjuanensis (Tetrapoda: Batrachosauria) from the Lower Permian of Utah and New Mexico. PaleoBios 16 (4): 1-8.
Laurin, M. 1996a. A reappraisal of Utegenia, a Permo-Carboniferous seymouriamorph (Tetrapoda: Batrachosauria) from Kazakhstan. Journal of Vertebrate Paleontology 16 (3): 374-383.
Laurin M. 1996b. A reevaluation of Ariekanerpeton, a Lower Permian seymouriamorph (Tetrapoda: Seymouriamorpha) from Tadzhikistan. Journal of Vertebrate Paleontology 16: 653-665.
Laurin, M. & R. R. Reisz. 1997. A new perspective on tetrapod phylogeny. In S. Sumida and K. Martin (eds.) Amniotes Origins: Completing the Transition to Land: 9-59. London: Academic Press.
White, T. E. 1939. Osteology of Seymouria baylorensis Broili. Bulletin of the Museum of Comparative Zoology 85: 325-409.
Zhang, F., Y. Li, & X. Wan. 1984. A new occurrence of Permian seymouriamorphs in Xinjiang, China. Vertebrata PalAsiatica 22: 294-304.
Information on the Internet
Available translations:A Portuguese translation by Artur Weber for homeyou.com.
Title Illustrations
| Scientific Name | Seymouria baylorensis |
|---|---|
| Location | Texas |
| Reference | Reproduced from White, T. E. 1939. Osteology of Seymouria baylorensis Broili. Bulletin of the Museum of Comparative Zoology 85:325-409. |
| Specimen Condition | Fossil -- Period: Lower Permian |
| Copyright | © 1939 The President and Fellows of Harvard College |
About This Page
I wish to thank my wife (Ms. Patricia Lai), Mr. John Hutchinson, and Mr. Matthew Marlowe who edited this page. I am indebted to Dr. David Maddison who provided invaluable help in linking this page with other pages on the Tree of Life. I thank Dr. Jozef Klembara for his useful comments on Discosauriscus.
Michel Laurin

Muséum National d'Histoire Naturelle, Paris, France
Correspondence regarding this page should be directed to Michel Laurin at
Page copyright © 1996 Michel Laurin
 Page: Tree of Life
Seymouriamorpha.
Authored by
Michel Laurin.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Seymouriamorpha.
Authored by
Michel Laurin.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- Content changed 15 January 1998
Citing this page:
Laurin, Michel. 1998. Seymouriamorpha. Version 15 January 1998. http://tolweb.org/Seymouriamorpha/15005/1998.01.15 in The Tree of Life Web Project, http://tolweb.org/